Here is the Biology First year book 2025, 1st Chapter from the Punjab Textbook Board, according to the new syllabus.
Read new books
| Chemistry book 2025 new syllabus. | Read |
| Computer Science book 2025 new syllabus. | Read |
| Biology book 2025 new syllabus. | Read |
| Islamiat book 2025 new syllabus. | Read |
| English book 2025 new syllabus. | Read |
| Physics book 2025 new syllabus. | Read |
| Urdu book 2025 new syllabus. | Read |
Chapters are available in full-screen mode with controls.
Introduction
The cell
Cells are the building blocks of all life, the smallest units that perform essential tasks like energy production, growth, reproduction, and maintaining balance within organisms. Understanding cells is key to grasping how life works, from simple bacteria to complex humans. In this chapter, we will explore the basic principles of cell theory, examine the different parts of cells, and learn about the processes that keep them functioning. We will also look at the microscopes used to study cells and the potential of stem cells in medicine. This chapter will help you understand why cells are essential in biology and how they support all life forms.
: Cell: Basic Structural and Functional Unit of Life
Cells are the basic building blocks of life, representing essential life characteristics in their microscopic structure. As the smallest units capable of life, they exhibit seven key properties: movement, respiration, homeostasis, growth, reproduction, excretion, and nutrition. These traits distinguish living organisms from non-living matter, offering insights into life's diversity at the cellular level.
Understanding how each of these properties is manifested at the cellular level provides insights into the complexity and functionality of life.
Movement: Cells exhibit internal movements, such as the movement of organelles and transport of substances across the cell membrane. Some specialized cells like white blood cells can also move through their environments using mechanisms like amoeboid movement.
Respiration: Cellular respiration involves converting glucose into ATP. This process is vital for maintaining the cell's activities and survival.
Homeostasis: Cells maintain stable intimal conditions through mechanisms like selective permeability of membranes and regulatory pathways, despite external environmental changes. Growth: Cellular growth includes an increase in cell size and the accumulation of mass, often as precursor to cell division.
Reproduction: Cells reproduce by dividing, either through mitosis for somatic cells, resulting in two identical daughter cells, or meiosis for gametes, contributing to genetic diversity
Excretion: Cells eliminate waste from cellular activities, including metabolic byproducts like carbon dioxide and urea, to maintain a healthy cellular environment.
Nutrition: Cells take in nutrients like oxygen, water and organic molecules from their environment for cellular functions. Autotrophic cells can produce nutrients via photosynthesis, while heterotrophic cells absorb and digest external nutrients. Each of these properties contributes to the overall functionality of cells and is essential for the sustenance and continuation of life.
The cell theory, a fundamental concept in biology, emerged from the advancements in microscopy in the 17th century. Pioneers like Robert Hooke, who first used the tenn "cell" in 1665 after observing cork under a microscope, and Antonie van Leeuwenhoek, who described live cells in pond water, laid the groundwork for this theory. In the 19th century,
Matthias Schleiden and Theodor Schwann significantly advanced the theory by proposing that all plants and animals are composed of cells, leading to the establishment of the cell theory's three main principles, given below.
Principles of the Cell Theory: The cell theory traditionally includes three basic principles:
All living organisms are composed of cells. They are the basic structural and functional units of life.
The cell is the smallest unit of life. It is the smallest structure capable of performing all the functions necessary for life.
All cells arise from pre-existing cells. This principle opposes the earlier notion spontaneous generation and emphasizes. the continuity of life.
The validation of cell theory historically involved several key experiments and observations:
Experiments that isolate single cells in culture demonstrate that individual cells can survive and function independently, maintaining homeostasis and responding to their environment.
Louis Pasteur's experiments that disproved spontaneous generation supported the idea that all
cells come from existing cells, not from non-living matter.
Modem techniques in imaging and molecular biology have provided detailed insights into DNA replication, mitosis and cytokinesis, conclusively showing that cells originate solely through the division of pre-existing cells.
While the cell theory provides a foundation for understanding biological structure and function,
there are exceptions:
Often debated whether they are "alive", viruses are not made up of cells but are genetic material (DNA or RNA) enclosed in a protein coat. They require a host cell to replicate, challenging the definition of cells as the smallest unit of life.
Like viruses, prions and viroid are both exceptions to cell theory, lacking cellular structure. Both challenge traditional views of life as organized in cellular structures.
The lack of a nucleus in red blood cells (RBCs) and sieve tube elements, which inhibit their ability to reproduce, stands as a notable exception to the cell theory.
The mycelium of some fungi consists of a network of hyphae that form a multicellular structure without fully divided cellular sections, creating large, multinucleated cells that can span significant distances, challenging the notion that all organisms are made up of discrete, individual cells.
: Microscopy
Microscopy is the science and technology of using microscopes to observe objects that cannot be seen with the naked eye- It is crucial in various scientific fields, including biology, medicine and nanotechnology. There are many microscopes, each with a different working principle However, magnification and resolution are fundamental concepts considered in designing and operating all types of microscopes. Magnification refers to the degree to which an object is enlarged when viewed through a microscope. It indicates how many times larger the object appears compared to its actual size.
Resolution is the ability of a microscope to distinguish between two closely spaced objects. It is determined by the wavelength of the light or electrons used in microscopy. Higher resolution allows for more precise and more detailed images.
In this topic, we will discuss only light microscope and electron microscope along with their working principle.
A light microscope is termed a compound microscope because it utilizes a system of multiple lenses, specifically objective lenses and an ocular lens, to magnify the objects (see Figure 1.1)- This configuration allows for significant magnification and detailed observation This multi-lens setup distinguishes it from simpler, single-lens microscopes
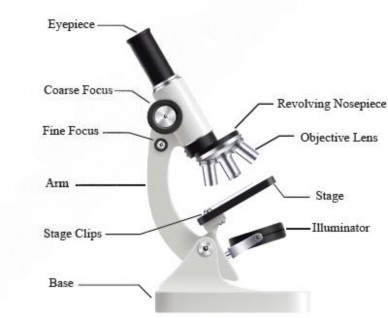
Figure 1.1: Parts of a light microscope
Working principle: A light microscope uses visible light to illuminate a specimen, which then passes through an objective lens present in the revolving nosepiece close to the specimen for initial magnification- This magnified image is further enlarged by an eyepiece or ocular lens present in the eyepiece (see Figure 1.2). By adjusting the objective lens's distance from the specimen, the image can be focused. The light, either transmitted or absorbed by the specimen, creates a contrasted image that allows for detailed observation of tiny structures.
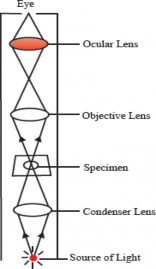

Figure 1.2: Principle of compound light Figure 1.3: Electron microscope
Magnification and Resolution: To calculate the total magnification, simply multiply the magnification power (4x, lox, 40x and 100x) of the objective lens by the magnification power (usually lox) of the ocular lens. For example, If you are using an objective lens with a magnification of 40x and an ocular lens with a magnification of lox, the total
magnification would be: 40xX 10x=400x
This means the image is magnified 400 times larger than the actual size of the object. Though it has magnification up to 1500x, it is less capable of distinguishing very fine details. Limited by visible light's wavelength, its resolution is about 200 nanometers. Light microscopes can be used to observe both live and dead samples. Ideal for live samples, it observes living cells and tissues in their natural state with minimal preparation, often using stains for enhanced contrast. Knowledge Booster The resolution of the human naked eye, in terms of distinguishing between two points or objects, is typically about 0.1 mm at a distance of about 25 cm from the eye.
Invented in the 1930s, electron microscopes use electron beams instead of light to illuminate samples. This technology provides a much deeper look into the fine details and surface feature of cells, including some of the larger molecules inside them (see Figure 1.3).
Working principle: An electron microscope operates by using a beam of electrons emitted from a heated filament in an electron gun. These electrons are accelerated and focused onto the specimen with electromagnetic lenses (see Figure 1.4). In a Transmission Electron Microscope (TEM), electrons passing through the very thin section of the specimen create an image on a photographic plate based on how they are absorbed and scattered. In a Scanning Electron Microscope (SEM), the image is formed from electrons scattered back from the specimen surface, producing a detailed 3D topographic image. The entire system is maintained under vacuum to prevent electron scattering by air, ensuing high-resolution imaging.
Magnification and Resolution: Electron microscopes have a much higher resolution, typically around O_ I nm for transmission electron microscopes (TEM), allowing for the visualization of structures at the molecular level. It can magnify up to 50 million times, offering detailed views of a specimen's ultrastructure. Only dead, fixed samples can be observed because the preparation process and the vacuum required for operation make it impossible to view live specimens- Samples must also be thinly sliced and sometimes stained with heavy metals for enhanced contrast.
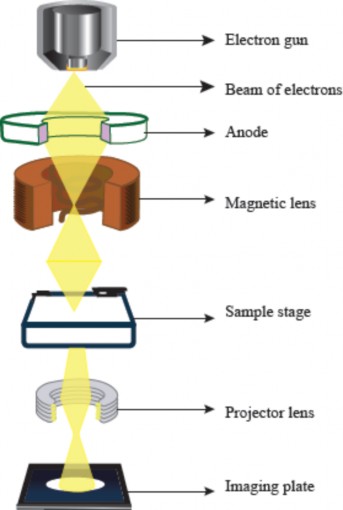
Figure 1.4: Working principle of transmission elctron microscope
: Ultrastructure of Animal and Plant Cells
We have studied the ultrastructure of animal and plant cells in grade 9 and will discuss it more in detail in this chapter There are cellular structures that are common in both animal and plant cells namely; plasma membrane, nucleus, cytoplasm, mitochondria, endoplasmic reticulum, Golgi apparatus and ribosomes, etc. However, both cells have umque organelles that are exceptional to one cell, for example, lysosomes and centrioles to animal cells and cell wall, chloroplast, large central vacuole and chloroplast to plant cells (see Figure I -5 & 1.6).
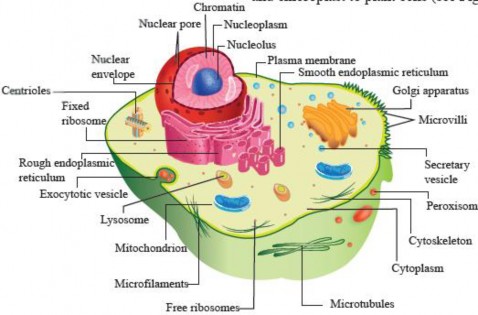
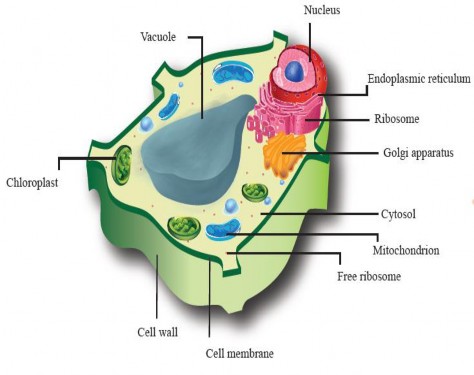
Figure 1.5: Ultrastructure of an animal cell.
Figure 1.6: Detailed structure of a plant cell.
These structures are integral to the function of both types of cells and illustrate the unique characteristics and adaptations of animal and plant cells to their respective environments and roles within organisms.
Structure and Functions of sub-cellular Organelles
A typical eukaryotic cell consists of the following components:
Cell wall and plasma membrane
Nucleus
Cytoplasm with its embedded organelles.
Cell Wall
The cell wall is an extracellular structure located outside the plasma membrane in plant cells. This is one of the features that distinguishes plant cells from animal cells. The cell wall provides support, shape, and protection against invading bacteria and fungi- It also helps maintain turgor pressure, which is essential for keeping the plant upright and rigid Plant cell walls are composed of cellulose fibers embedded in a network of carbohydrates. The structural arrangement of cellulose and other polysaccharides results in a porous matrix that is rigid enough to provide structural support but porous enough to allow the movement of molecules through it. The plant cell wall has three main layers as shown in Figure 1.7. The initial cell wall formed by all plant cells is the primary cell wall, which is relatively thin and flexible. As the cell grows, the primary cell wall increases m size. It is composed of cellulose microfibrils embedded in a matrix of hemicellulose, pectin, and proteins. The secondary cell wall is much thicker and more rigid than the primary wall and is deposited inside the primary wall once the cell has stopped expanding. It is lich in cellulose, hemicellulose, inorganic salts, waxes and often lignin, which provides additional strength and resistance. Not all plant cells contain a secondary cell wall. The formation of the seconda1Y cell wall occurs in specific cell types, such as xylem cells and sclerenchyma cells- These cells are typically involved in providing structural support and rigidity to the plant.

Figure 1.7: Structure of a plant cell
A layer of gel-like polysaccharides that binds the primary cell walls of adjacent cells together is called the middle lamella. It is mainly composed of pectins, which are complex polysaccharides rich in galacturonic acid. Pectins are highly hydrophilic (water-loving) and gelatinous, allowing them to fill spaces between cells effectively. Additionally, calcium and magnesium ions interact with the pectin molecules, increasing the rigidity and stability of the middle lamella. Both primary and secondary cell walls have minute channels or perforations called plasmodesmata. These allow ions and small molecules to move directly from one cell to another through the connecting cytosol, bypassing the plasma membranes or cell walls.
Cell walls also surround the cells of fungi, some protists, and prokaryotes. Bacterial cell walls are composed of peptidoglycan, whereas those of fungi contain chitin. As mentioned earlier, animal cells do not form rigid, external cell walls. However, most animal cells secrete extracellular material and have other structures at the cell surface that play vital roles in the support and protection of cells
Cell Membrane
The plasma membrane, also known as the cell surface membrane, is a fundamental feature of both eukaryotic and prokaryotic cells. It is an extremely thin structure, measuring approximately 5-10 nm in thickness, yet it is strong enough to preserve the integrity of the cell. This membrane not only encases the cell’s internal components but also acts as a selective barrier, regulating the passage of substances into and out of the cell.
Structure of cell membrane: The Fluid Mosaic Model, proposed by S.J. Singer and Garth Nicolson in 1972, revolutionized our understanding of the cell membrane's structure. This model highlights the fluid nature of the lipid bilayer and describes the
mosaic-like arrangement of proteins that are embedded within it The cell membrane contains collection of various components, including lipids, proteins, and carbohydrates.
The foundation of the cell membrane is the phospholipid bilayer, made up of two layers of phospholipids- Each phospholipid molecule has a hydrophilic head (water-attracting) and two hydrophobic tails (water-repelling). Proteins embedded within this lipid bilayer include integral proteins, often spanning its entire width, and peripheral proteins, which attach to its surfaces. Cholesterol is also interspersed within the lipid bilayer.
Carbohydrates, in the form of short-chain polysaccharides, are present on the cell surface membrane- Some are attached to proteins forming glycoproteins and others to lipids forming glycolipids, located only on the outer surface of the membrane (see Figure 1.8)
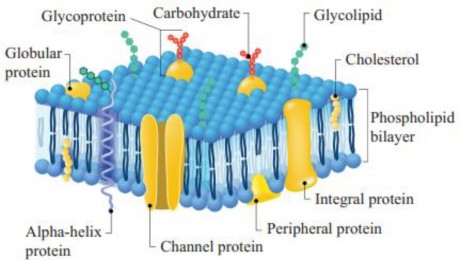
Figure 1.8: Detailed structure of plasma membrane
Role of different components of cell membrane:
The phospholipid bilayer acts as a selective barrier to most water-soluble substances. The non- polar nature of the phospholipid tails form a hydrophobic core that prevents the passage of polar molecules such as sugars, amino acids, and proteins. This barrier function helps to maintain cellular integrity by preventing the leakage of these water-soluble molecules out of the cell and barring entry to unwanted substances.
Cholesterol plays a crucial role in maintaining the membrane's fluidity and stability. It increases fluidity at low temperatures, preventing rigidity by avoiding close packing of phospholipid tails. This adaptation allows cells to survive colder temperatures. Similarly, cholesterol helps stabilize the membrane at higher temperatures, preventing it from becoming too fluid and ensuring mechanical stability. Without it, membranes can easily break, causing cells to burst. Proteins in the membrane serve various functions: those which are involved in the transport of substances are called channel proteins and those which combine with substances to move them across the membranes are called carrier proteins. Others act as enzymes, catalyzing reactions the membrane's surface. Some proteins participate in signal transduction by acting as receptors that
can receive and respond to various external signals, such as hormones and neurotransmitters (see Figure 1.9).
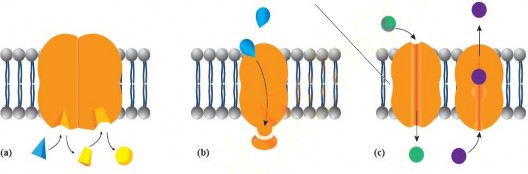
Figure 1.9: (a) Enzyme activity (b) Protein as a receptor in signal transduction (c) Protein as a channel or carrier in the transport of molecule.
Moreover, glycolipids and glycoproteins serve as cell markers, facilitating cell to cell recognition. Additionally, some glycoproteins are involved in cell adhesion, enabling cells to interact and bond with each other, which is fundamental for the formation of tissues and organs.
The cell membrane's structure and functions are integral to the survival and proper functioning of cells in organisms, serving as a dynamic interface that allows cells to interact with their surroundings while maintaining internal stability.
Phospholipids: Phospholipids in cell membrane are derivatives of phosphatidic acid, containing a backbone of a glycerol molecule (three carbon compounds with an -OH group on each carbon). Two fatty acid chains are attached to the first and second carbons of the glycerol molecule (see Figure 1.10). The chain attached to the first carbon is saturated, and the one attached to the second carbon is unsaturated. A phosphate group, along with other organic compounds (see Chapter 2), forms the polar head of the phospholipid attached to the third carbon of glycerol.

Figure 1.10: Chemical structure of a phospholipids molecule
Techniques Used to Study the Cell Membrane:
Various techniques are in use to study the structure and function of the cell membrane.
Electron Microscopy: Transmission and scanning electron microscopy, provides detailed images of cell membranes and their fine structure at high magnifications.
Fluorescence Microscopy: Fluorescent dyes or proteins are used to label specific components of the cell membrane, too visualize and track membrane proteins or lipids in living cells.
X-Rays Crystallography: This technique is used to determine the 3D structures of membrane proteins, providing detailed information about their shapes and interactions within the lipid bilayer.
Fracture Electron Microscopy: Freezing and fracturing of cells to reveal the internal structures of the membrane, including the arrangement of proteins and lipids.
Cell Fractionation: Fractionation techniques isolate and purify cellular components, including membranes, for biochemical analysis, allowing researchers to study the composition and functions of membrane proteins and lipids.
Membrane Transport mechanisms
Cells require a constant supply of nutrients from their environment to carry out their metabolic functions. Nutrients such as glucose and amino acids must be transported into the cell to fuel these metabolic processes. Conversely, as cells metabolize these nutrients, they generate waste products like carbon dioxide and ammonia, which, if accumulated, could be toxic. However, the cell membrane does not let just anything pass through because it is selectively permeable. To manage this, cells use various transport mechanisms to intake these substances into the cell and expel the wastes, maintaining a safe internal environment. The following are the mechanisms responsible for the transport of substances through the cell membrane:
Simple Diffusion: Simple diffusion is a process by which molecules move from an area of higher concentration to an area of lower concentration across a cell membrane without the need for energy (passive transport). This process can occur only if the membrane is permeable to the substance concerned. The lipid bilayer is permeable to lipid soluble compounds such as steroids and fatty acids, as well as small molecules like oxygen and carbon dioxide, which diffuse through it rapidly. Diffusion begins with a concentration gradient, a difference in the concentration of a substance between two areas. Molecules, driven by their kinetic energy, spread out into available space. This movement continues until the concentration on both sides of the membrane equalizes, reaching a state of dynamic equilibrium (see Figure 1.11). At equilibrium, molecules continue to move, but there is no net movement in either direction. It is also noteworthy that different particles can diffuse in opposite directions simultaneously without interference. For example, carbon dioxide (CO2) produced as a by-product of cellular respiration diffuses out of the cells into the surrounding tissues, while O diffuses into these cells to support cellular respiration.
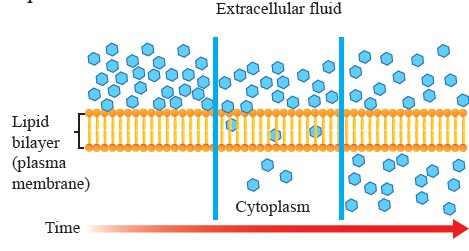
Figure 1.11: Diffusion through a permeable membrane.
The rate of simple diffusion is influenced by factors such as the concentration gradient, temperature (higher temperatures increase molecular movement), molecular size (smaller molecules diffuse more quickly), and the permeability of the membrane.
Facilitated Diffusion: Substances like polar molecules, including glucose and amino acids, and ions like sodium (Na+), potassium (K+) and chloride (CD cannot diffuse through the phospholipid bilayer They need the help of specific protein molecules to cross the membrane. Facilitated diffusion is the process that allows these particular substances to move across cell membranes from an area of higher concentration to an area of lower concentration, using transport proteins embedded in the membrane, without the expenditure of energy.
There are two main types of transport proteins involved in this process: channel proteins and transporter/carrier proteins. Channel proteins form passageways or pores that enable specific molecules to cross the membrane. Some of these proteins are always open, providing a constant pathway, while others are gated and can be opened or closed as needed (see Figure 1.12 a&b). Transporter proteins, in contrast, are more selective. They bind to a specific molecule or ion on one side of the membrane and then transport these molecules to the other side without forming an open channel. For example, glucose enters a cell through a glucose transporter. Unlike channel proteins, transporter proteins only open to one side at a me (see Figure
1.12 c).
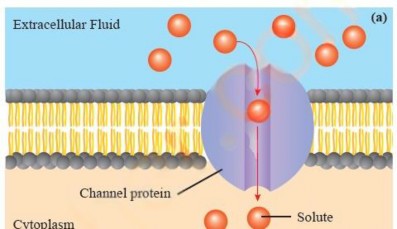

(b)
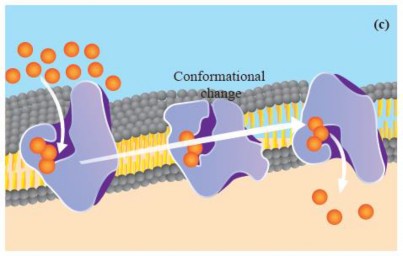
Figure 1.12: Faciliated diffusion across the plasma membrane. (a) Open channel protein (b) Gated channel protein (c) Carrier protein
Like simple diffusion, facilitated diffusion occurs down the concentration gradient. However, it does so at a faster rate than simple diffusion for specific molecules.
Osmosis: Osmosis is a special type of diffusion that involves the passive movement of water molecules through a selectively permeable membrane from an area of higher water concentration (lower solute concentration) to an area of lower water concentration (higher solute concentration) as show in Figure I. 13.
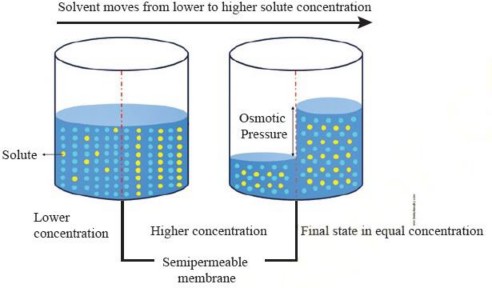
Figure 1.13: Osmosis
Direction of Osmosis: The net direction of osmosis depends on the difference in solute concentration on either side of the membrane (Study Table 1.1). When the solute concentration outside the cell is lower than inside, the external solution is termed hypotonic relative to the inside (cytosol). Under these conditions, water moves into the cell until equilibrium is achieved. Conversely, when the solute concentration outside is higher than inside, the external solution is considered hypertonic to the cytosol. In this case, water moves out of the cell until equilibrium is reached.
Condition | Net movement of water | |
External solution s hypotonic to cytosol | Into the cell |
|
Extremal solution is hypertonic to cytosol | Out of the cell |
|
Extremal solution is isotonic to cytosol | None |
|
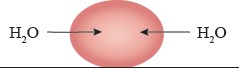

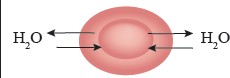
However, when the solute concentration is the same inside and outside the cell, the external solution is called isotonic to the cytosol. In this situation, water moves into and out of the cell at the same rate, so there is no net movement of water.
Active Transport: If a cell depended exclusively on diffusion for the exchange of substances, it would be unable to regulate its internal environment effectively. This means that any substance with a higher concentration outside the cell could enter, regardless of its necessity or potential harm, while essential substances might leave the cell whenever their concentration inside exceeded that on the outside. To address these challenges, the cell membrane employs a mechanism known as active transport. This process enables the molecules or ions to move against their concentration gradient, from an area of lower concentration to an area of higher concentration, with expenditure of energy. Active transport relies on specialized transmembrane proteins called pumps or transporters. Following are the two types of active transport:
Primary active transport: Primary active transport directly uses ATP to fuel the transport of molecules. One common example of primary active transport is the ion pump, such as the sodium-potassium pump which actively transports three sodium ions (Na+) out of the cell and two potassium ions (K+) into the cell, using ATP to power the process (see Figure l. 14).
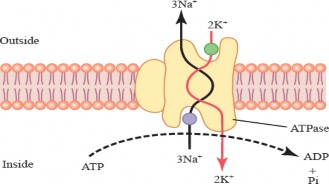
Figure 1.14: Primary active transport through sodium-potassium pump.
Secondary Active Transport (Co-transport): Secondary active transport utilizes the energy released from the movement of one molecule down its concentration gradient to drive the movement of another molecule against its gradient. This process does not directly use ATP; instead, it relies on the energy gradient created by primary active transport. For example, the sodium-glucose transport protein (SGLT) uses the sodium gradient established by the Na+/K+ pump to transport glucose into cells (see Figure 1.15).

Figure 1.15: Primary and secondary active transport
Endocytosis and Exocytosis: Macromolecules such as proteins and polysaccharides are often too large to be moved through the cell membrane by transport proteins. Endocytosis and exocytosis are essential cellular processes that transport these large molecules and panicles across the cell membrane by using energy.
Endocytosis: Endocytosis is the process by which cells take in substances from the extemal environment by engulfing them into a vesicle formed from the cell membrane. Phagocytosis and pinocytosis are the two main forms of endocytosis.
Phagocytosis: In phagocytosis, solid panicles are engulfed by the cell. This process begins when the phagocyte identifies a target particle and extends pseudopodia (false feet) to surround and enclose it within a vesicles or phagosome [see Fig 1.16 (a)]. This mechanism is common among multicellular organisms like amoeba and also occurs in humans. Certain white blood cells in humans are capable of engulfing debris like worn-out red blood cells or bacteria.
Pinocytosis: In pinocytosis, small droplets of fluid or dissolved substances are engulfed by the cell. The cell membrane invaginates, creating small vesicles called pinosomes that contain the ingested molecules [see Figure 1.16 (b)]. Microvilli, bristle-like protrusions in the small intestine, use pinocytosis to absorb nutrients (amino acids, vitamins, etc.) from food.
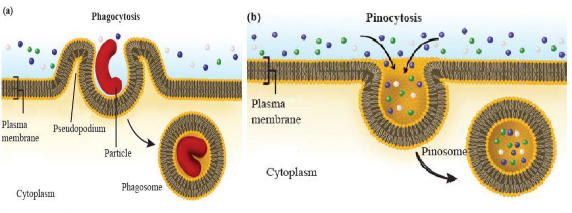
Figure 1.16: Types of endocytosis (a) Phagocytoss (b) Pinocytosis
Exocytosis: Exocytosis is the process by which cells expel substances from the cell interior to the extracellular space. This involves the fusion of vesicles containing cellular products with the cell membrane, thereby releasing the contents outside the cell (see Figure 1.17). An example of this is the secretion of digestive enzymes from pancreatic cells, where secretory vesicles from the Golgi apparatus carry enzymes to the cell surface for release.
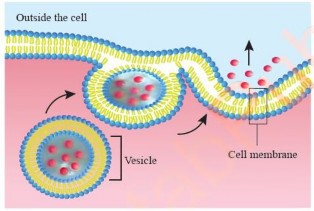
Figure 1.17: Exocytosis
The nucleus is a prominent organelle in eukaryotic cells, responsible for storing the cell's genetic material and acting as its control center. It is typically spherical or oval and has an average diameter of 5 gm. The nucleus is composed of the following key components, each with its important role to play:
The nuclear envelope, also known as the nuclear membrane, is a double-layered barrier that surrounds the nucleus, effectively separating it from the cytoplasm. The outer layer of the nuclear envelope is continuous with the endoplasmic reticulum (ER) membrane, while the inner layer faces the nucleoplasm. The space between the inner and outer membranes is called the perinuclear space. The nuclear envelope is perforated with nuclear pores, which are protein channels spanning both membranes (see Figure 1.18). These pores allow for the passage of materials between the cytosol and the nucleus. Each nuclear pore is made up of specialized proteins called nucleoporins.

Figure 1.18: Structure of Nucleus
Within the nucleus lies the nucleoplasm, a gel-like substance, which is primarily made up of water, dissolved ions such as K+, Ca++ and Mg++, nucleotides, enzymes, RNA and proteins (histone and non-histone proteins). It also contains chromatin, which is formed by wrapping DNA around histone proteins, as well as the nucleolus and other nuclear components. Chromatin stores genetic information in a loosely coiled state during the non-dividing stage of the cell. During cell division, chromatin condenses into visible chromosomes.
The nucleolus is a prominent structure within the non-dividing nucleus. Is a dense, It non- membrane-bound region and plays a key role in ribosome biogenesis. The nucleolus comprises a central fibrillar area surrounded by a peripheral granular area. The central fibrillar area is primarily involved in the transcription of rRNA (ribosomal RNA) from rDNA, while the peripheral granular area is associated with the formation of both the small and large subunits of ribosomes. These subunits are then transported to the cytoplasm through nuclear pores, where they join to form functional ribosomes.
Cytoplasm is the living component and lies between the nuclear membrane and cell membrane. It is gel-like in nature and is composed of water (about 90%) and embedded substances. The embedded substances include the organelles and chemicals consisting of ions, molecules like salts, vitamins and dissolved gases. The water and suspended or dissolved substances form the cytoplasmic matrix. The dissolved micromolecules make it a true solution, while the macromolecules, such as proteins, make it a colloidal solution. Cytoplasm can be differentiated into two parts;
Ectoplasm: The outer or the peripheral portion of the cytoplasm is comparatively non granular, viscous and gel-like (Cytogel).
Endoplasm: The inner or central portion of the cytoplasm is granular, contains organelles and is less viscous (Cytosol).
The cytoplasm acts as a site for certain metabolic reactions, e.g. glycolysis, protein synthesis and mitosis. It is a reservoir of important chemicals such as hormone and enzymes. Cellular wastes are also dissolved here. Cytoplasm houses a number of cell organelles that perform specific function in the cell. The cell organelles found in eukaryotic cells are Ribosomes, Endoplasmic Reticulum, Golgi bodies, Lysosomes, Peroxisomes & Glyoxysomes, Centriole, Mitochondria and Plastids.
Ribosomes are cellular structure which are involved in protein synthesis in living cells. Ribosomes are ribonucleoprotein complexes that are composed of ribosomal RNA (rRNA) and proteins. They consist of two subunits, the large and the small subunits, and are found in both prokaryotic and eukaryotic cells (see Figure 1.19). Prokaryotic ribosomes are typically smaller and measure about 70S, while eukaryotic ribosomes are slightly larger and measure about 80S.

Figure 1.19: Large and smaller subunit of ribosome
In both prokaryotes and eukaryotes, these small and large subunits join to form a functional ribosome only when attached to an mRNA molecule. Ribosome catalyzes the formation of peptide bonds between amino acids during proteins synthesis. The small subunit reads the mRNA sequence, while the large subunit helps to link the amino acids together to form the growing polypeptide chain. However, when a cell has a high demand for protein synthesis, such as during cell growth, replication, and division, multiple ribosomes translate a mRNA simultaneously, forming a complex called polysomes or polyribosomes (see Figure 1.20). This structure significantly increases the efficiency of protein synthesis.
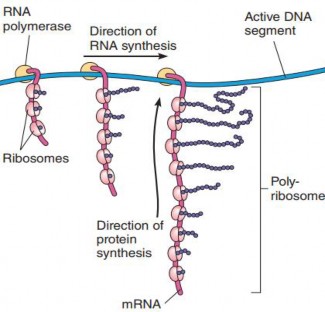
Figure 1.20: Polysome formation
Eukaryotic ribosomes may be either free, or bound to the rough endoplasmic reticulum. However, bound and free ribosomes are structurally identical. The number of ribosomes can vary depending on the cell type and its specific needs. Cells that have high rates of protein synthesis have a large number of ribosomes, as well as prominent nucleoli, given the role of nucleoli in ribosome assembly.
The endoplasmic reticulum is a vast network of channels and interconnected that spreads throughout the cell. It is comprised of membranous tubules and sacs, known as cisternae, and is continuous with the outer membrane of the nuclear envelope. The space within these tubular and sac-like structures of the ER is called the ER lumen or the endoplasmic reticulum cisternal space, and it is entirely enclosed by the ER membrane, separating it from the cytosol of the cell. There are two distinct regions of the ER that differ in structure and function- These are rough endoplasmic reticulum (RER) and smooth endoplasmic reticulum (SER).
Rough endoplasmic reticulum: The rough endoplasmic reticulum (RER) appeals rough under a microscope due to the presence of ribosomes on its outer surface (see Figure 1.21)- The rough ER is characterized by flattened sacs or cistenae, often extending from the nuclear envelope and is abundant in cells actively producing proteins for export, such as plasma cells and pancreatic cells- The rough endoplasmic reticulum (ER) plays an essential role in the synthesis and assembly of proteins, including those that are exported from the cell, destined for other organelles, or become a part of a membrane. Polypeptides, which are intended for export to other organelles, enter the ER lumen directly Enzymes within the ER lumen modify and assemble proteins, including protein folding and adding carbohydrates to them. Once modified and assembled, the proteins are packaged into transport vesicles. These transport vesicles bud off at a specialized area of rough ER that lacks bound ribosomes called the transitional ER and transport the proteins to their respective destinations within the cell, such as other organelles or the cell surface, for secretion.
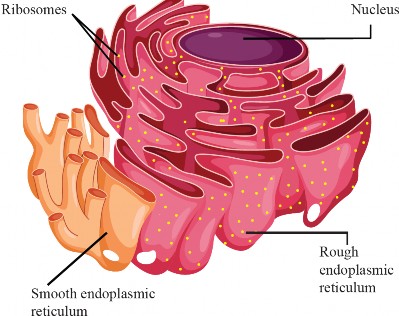
Figure 1.21: Endoplasmic reticulum
Smooth endoplasmic reticulum: The smooth ER, which lacks ribosomes on its surface, giving it a "smooth" appearance, has a tubular network structure. SER is involved in various functions, such as lipid metabolism, including the synthesis of lipids such as phospholipids and steroids. It also plays a role in the metabolism of carbohydrates and the detoxification of drugs and toxins by modifying them to be more water-soluble for easier removal from the cell- One of the most important functions of the smooth ER in animal cells is the production of sex hormones such as estrogen and testosterone Additionally, in muscle cells, a specialized type of smooth ER called the sarcoplasmic reticulum stores and delivers the calcium ions required for muscle contraction.
Both rough and smooth ER contribute to the biosynthesis of new cellular membranes, providing components like phospholipids and proteins necessary for membrane formation and growth.
The Golgi apparatus, also known as the Golgi complex or Golgi body, was named after its discoverer, Camillo Golgi. It is made up of a stack of slightly curved, flat, membrane-enclosed sacs called cisternae (sing. cisterna). Each cisterna has an internal space or lumen. The Golgi apparatus is divided into three distinct regions: the cis face, the trans face, and a medial region in between.
The cis face, which is the entry surface or forming face, is convex and located adjacent to the nucleus or transitional ER. It receives transport vesicles bringing molecules from the ER The trans face, which is the exit surface or maturation face, is concave and closest to the plasma membrane- It packages molecules in vesicles and transports them out of the Golgi.
The Golgi apparatus plays a key role in the modification and processing of proteins and in directing them to their proper destinations- Secretory proteins that are assembled in the rough ER lumen are first transported to the cis face of the Golgi complex via transport vesicles formed from the ER membrane- These proteins then progress through the Golgi stack, undergoing successive modifications until they reach the trans face of the Golgi, where they are packaged into secretory vesicles and shipped to their destination. Outgoing secretory vesicles may be used within the cell or proceed to the plasma membrane, where they discharge their contents during secretion. In animal cells, some of the vesicles that leave the Golgi contribute to the formation of lysosomes. The Golgi body contains enzymes that are particularly involved in the modification and formation of conjugated molecules such as glycoprotein and lipoprotein (see Figure 1.22).
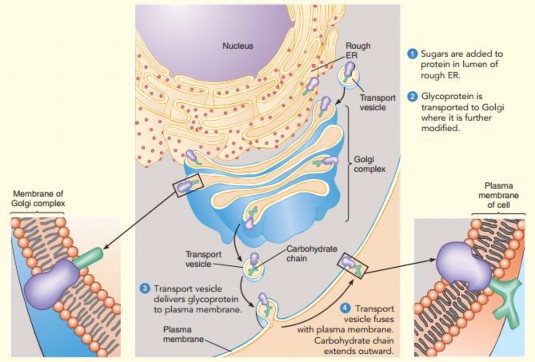
Figure 1.22: Protein synthesized at RER, are modified in the Golgi Apparatus and released as secretory vesicle
In plants, the Golgi body is mainly involved in the secretion of materials of primary and secondary cell walls. During cytokinesis of mitosis or meiosis, the vesicles originated from the Golgi apparatus coalesce in the phragmoplast to form a cell plate.
Lysosomes, derived from Greek ("lyso" meaning digestive and "soma" meaning body), are small vesicles filled with about 40 different types of digestive enzymes, such as lipases, nucleases and proteases, that are active under acidic conditions (pH around 5) as shown in Figure [1.23(a)]. An ATP-driven proton (H+) pump within the membrane of the lysosome maintains this acidic environment. If lysosomes break, open or leak, their enzymes become inactive due to the near- neutral pH (about 7.2) of the cytosol. However, extensive leakage from many lysosomes can lead to cell destruction through self-digestion.
The hydrolytic enzymes of lysosomes are synthesized in the rough ER and then transported to the Golgi apparatus. There, these enzymes are modified, and packaged into Golgi vesicles, which eventually bud off as primary lysosomes. Lysosomes facilitate both intracellular and extracellular digestion. In the case of intracellular digestion, the primary lysosome fuses with an incoming vesicle, such as a pinosome or phagosome, carrying food content from outside the cell via endocytosis to form a large vesicle called a secondary lysosome. Powerful enzymes in the secondary lysosome come in contact with the ingested molecules and begin the digestion process. Digestion products such as simple sugars, amino acids, and other monomers pass into the cytosol and become nutrients for the cell. In addition, certain human cells, like macrophages, help defend the body by engulfing and destroying bacteria and other invaders with the help of lysosomes.
Lysosomes also use their hydrolytic enzymes to recycle the cell's own material, which is called autophagy. During autophagy, the lysosome appears to flow around the damaged organelle and fuse to form autophagosome [see Figure 1.23(b)].
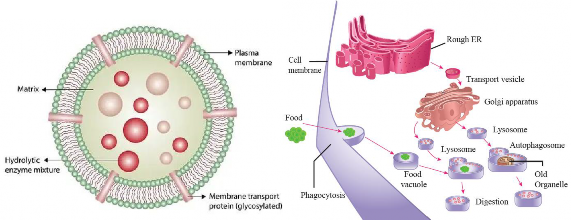
(a) (b)
Figure 1.23: (a) Structure of Lysosome (b) Formation and function od Lysosomes
The lysosomal enzymes dismantle the enclosed material, and the resulting small organic compounds are released into the cytosol for reuse. In this way, a cell continually renews itself.
In certain pathological conditions, when a cell dies, the lysosomal enzymes are released into the cell cytoplasm and begin to break down cellular components. This process is called autolysis. This can help in clearing dead cells and tissues.
Extracellular digestion by lysosomes is less common than intracellular digestion. It is a process where lysosomes release their hydrolytic enzymes outside the cell to break down substances in the extracellular matrix. For example, the heads of sperm contain a special lysosome called the acrosome, which digests a path to the ovum (egg) during fertilization. Similarly, during development, lysosomes facilitate cartilage replacement by breaking down the cartilage matrix, allowing for the formation of bone.
In certain genetic diseases of humans, known as lysosomal storage diseases, one of the digestive enzymes usually present in lysosomes is absent. For instance, In Tay-Sachs disease, a lipid- digesting enzyme is missing or inactive, and the brain becomes impaired by an accumulation of lipids in the cells. The accumulation of this lipid in brain cells causes severe intellectual disability, blindness, and death before age 4.
Microbodies are small, membrane-bound organelles present in nearly all eukaryotic cells, including those of plants, animals, fungi, and protists. Enclosed by a single lipid bilayer, they contain a collection of enzymes. A notable type of microbody is the peroxisome present in animal cells and higher plants, which houses oxidative enzymes such as oxidase, peroxidase, and catalase (see Figure 1.24).
Peroxisomes originate from the endoplasmic reticulum (ER), where specific proteins are integrated into ER-derived vesicles. These vesicles mature to form fully functional microbodies. Peroxisomes are especially abundant in cells involved in the synthesis, storage, or degradation of lipids. They play a crucial role in the oxidation of fatty acids, converting long-chain fatty acids into smaller molecules that the cell can use for energy production. This process generates hydrogen peroxide, a potentially harmful by product. However, peroxisomes are equipped with catalase, an enzyme that efficiently converts hydrogen peroxide into water and oxygen, thus neutralizing its toxicity. In liver and kidney cells, peroxisomes also detoxify various toxic compounds, including ethanol, converting them into less harmful products for elimination.

Figure 1.243: Structure of Peroxisome
Certain plant seeds, rich in fats and oils, such as corn, soybean, sunflower, and peanut, contain specialized peroxisomes called glyoxysomes. It contains enzymes that are involved in the glyoxylate cycle, which allows the conversion of stored lipids (fats) into sugars. The sugars are used by the young plant as an energy source and as a component for synthesizing other compounds.
A vacuole, like a vesicle, is a membrane-bound organelle found in the cells of plants, fungi, protists, and some animals. However, vacuoles are typically larger than vesicles. It is essentially a compartment filled with water that contains inorganic and organic molecules, including enzymes in solution. The membrane surrounding the vacuole is called the tonoplast.
Vacuoles are derived from the endoplasmic reticulum and Golgi apparatus. Immature plant cells contain numerous small vacuoles, which fuse together to form a large central vacuole as the cell matures (see Figure 1.25). The vacuole contains a high concentration of solutes and takes in water, creating hydrostatic pressure called turgor pressure, which helps plants stay rigid and upright. Plant cells grow by adding water to the central vacuole, which can occupy as much as 80% of the cell's volume. The vacuole also serves as a storage compartment for many compounds, including salts, sugars, and weak acids. Plant vacuoles contain hydrolytic enzymes and behave like lysosomes, breaking down wastes, damaged organelles, and other cell components. In some plant vacuoles, compounds that are poisonous to herbivores are stored as a means of defense.
In unicellular protists such as protozoa, food vacuoles (originate from the cell membrane by endocytosis) are present, which fuse with lysosomes that digest the food. Some protozoa also have contractile vacuoles, which remove excess water from the cell.

Figure 1.25: A large central vacuole in Plant cell
Vesicles (from the Latin word " vesicula," meaning "small bladder") are small sacs enclosed by lipid bilayers. These structures are essential for various cellular processes sac as transport, storage, an communication. Because of the phospholipids in their membranes, vesicles can bud off and fuse with other membrane structures within the cell (see Figure I .26).
Vesicles come in different sizes and serve specialized functions. They are involved in transporting molecules within the cell. For example, proteins from the endoplasmic reticulum are transported to the Golgi apparatus through transport vesicles. Additionally, secretory vesicles originating from the Golgi apparatus can release their contents outside the cell by fusing with the cell membrane, a process vital for the secretion of various substances.

Some vesicles, called endocytic vesicles, are formed during endocytosis, (where they engulf external materials and bring them into the cell for processing. Another example of a vesicle, lysosomes, contains digestive enzymes that break down waste materials and maintain cellular balance. Some vesicles are specialized for storage; for instance, synaptic vesicles in nerve cells store neurotransmitters that are released during synaptic transmission.
Figure 1.26: Transport vesicles transferring proteins from ER to Golgi apparatus
Centrioles are cylindrical structures that usually occur in pairs and are located near the nucleus in a region called the centrosome. Within the centrosome, a pair of centrioles lies at right angles to each other. Centrioles are known as "9 x 3 structures" because each centriole is composed of nine sets of microtubule triplets arranged to form a hollow cylinder (see Figure I .27). These microtubule triplets are surrounded by a protein matrix, which supports and stabilizes their structure. Centrioles are mainly found in animal cells and are absent in fungi and plant cells. During the S phase of cell division (mitosis and meiosis), a pair of centrioles duplicate and later migrate to opposite poles of the cell during Prophase. These centrioles serve as the primary organizing centers for the formation of spindle fibers (for details, see Knowledge 1.8), which are necessary for the separation of chromosomes into daughter cells. The complete set of spindle fibers and centrioles is collectively called mitotic apparatus. Centrioles are also involved in the formation of basal bodies of cilia and flagella, which are hair-like structures that project from the cell surface.
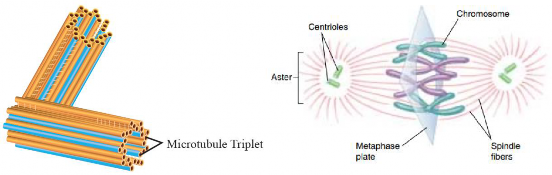
Figure 1.27: A pair of centrioles forming spindle fibers during cell divison
The term 'mitochondrion' is derived from the Greek words "mitos" and "chondrion" which means "thread" and "granules-like", respectively. They are often known as the "powerhouse of the cell" due to their crucial role in energy production. Mitochondria are the double membranous organelles that are found in most eukaryotic cells. The outer layer of mitochondria is smooth and encloses the entire organelle. It contains special proteins called porins that form channels that allow molecules such as ions, AT P, and other metabolites to pass through. The inner I membrane is folded into projections called cristae, which increase the surface area for chemical reactions. The inner membrane contains ATP synthase enzymes that are required for ATP production. ATP synthase enzymes consist of two components: Fo and F1 particle (see detail of ATP synthase in Chapter 10). The space between the outer and inner membranes is called inter-membranous space. The Inner membrane encloses the innermost compartment called the mitochondrial matrix (see Figure 1.28). It contains enzymes responsible for the Krebs cycle (citric acid cycle) and other metabolic reactions that occur in the matrix. It also contains mitochondrial DNA (mtDNA) and ribosomes for synthesizing some mitochondrial proteins.
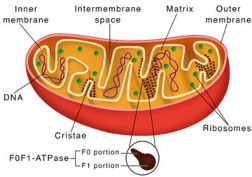
Figure 1.28: Structure of mitochondria
Mitochondria play a multifaceted role in cellular metabolism beyond ATP production. It has metabolic pathways for carbohydrates, fats, and proteins, converting these compounds into usable energy and other crucial molecules necessary for cellular processes. It is also involved in calcium regulation. In specialized brown fat cells, mitochondria contribute to heat generation, a process known as non-shivering thermogenesis.
Plastids are organelles found in the cells of plants and algae. There are three main types of plastids:
Chromoplasts are involved in pigment synthesis and storage other than chlorophyll, giving fruits, flowers, roots, and aging leaves their distinctive colours.
Leucoplasts are colourless plastids found in roots and seeds, where they store food. Chloroplasts contain green pigment chlorophyll and are involved in photosynthesis, the process by which light energy is converted into chemical energy (carbohydrate) using of carbon dioxide and water.
In plants, chloroplasts are primarily located within the mesophyll cells of the leaf, with each mesophyll cell containing 20 to 100 chloroplasts. Like mitochondria, chloroplasts are enclosed by outer and inner membranes, forming the chloroplast envelope. These membranes are separated by a very nanow intermembrane space. The inner membrane encloses a fluid-filled region called the stroma, which contains most of the enzymes required for carbohydrate synthesis. Similar to mitochondria, chloroplasts also contain their own DNA and ribosomes.
Suspended in the stroma is a third membrane system forming interconnected, flat, disc-like sacs known as thylakoids. The thylakoid membrane encloses an interior space called the thylakoid lumen. In some regions of the chloroplast, thylakoid sacs are stacked to form structures called grana (singular, granum), with some thylakoid membranes extending between grana, often called lamella (see Figure 1.29). Like the inner mitochondrial membrane, the thylakoid membrane is involved in ATP synthesis.
Thylakoid membranes house several types of pigments that absorb visible light, with different pigments absorbing various wavelengths. Chlorophyll, the primary photosynthetic pigment, mainly absorbs light in the blue and red regions of the visible spectrum. Energy absorbed from sunlight excites electrons within the chlorophyll molecules; these – electrons are used in light- dependent reactions to produce ATP and NADPH. The ATP and NADPH generated are then utilized in the light-independent reactions (Calvin Cycle) in the stroma to convert carbon dioxide into glucose.
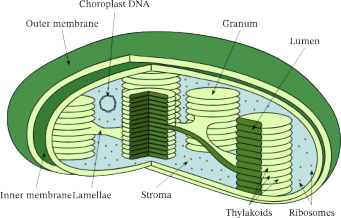
Figure 1.29: Structure of a chloroplast
: Comparison of Prokaryotic and Eukaryotic cell
Cells are classified into two types based on their structure, namely prokaryotic and eukaryotic and eukaryotic cells. Table 1.2 differentiate between these two cell types with diagrams.
FEATURES
PROKARYOTE CELLS
EUKARYOTE CELLS
NUCLEUS
The type of cell that lacks a nucleus is called a prokaryotic cell and the organism is called a prokaryote.
The type of cell that contains a well defined nucleus is called eukaryotic cell and the organism is called a
eukaryote
DNA
DNA is without any nuclear membrane
covering and is directly submerged in
cytoplasm-
DNA is enclosed inside the nucleus covered
by nuclear membrane.
MEMBRANE-BOUNDED STRUCTURES
Membrane-bounded absent
structures are
Membrane-bounded structures are present.
RIBOSOMES
They have •mall Gird 70S
ribosomes-
They have large sized 80S
ribosomes.
CELL WALL
Their cell wall is composed of polysacchande chains covalently
bonded with shorter chains of amino
acids forming peptidoglycan or murein.
Cell wall of plants is generally composed of cellulose while cell wall of fungi contains
chitin-
CELL DIVISION
They reproduce by binary fission.
They reproduce by mitosis and meiosis.
NUCLEOLUS
Nucleolus is absent.
Nucleolus is present.
CHOLESTEROL
Cholesterol is not found in the membrane
of prokaryotic cell-
Cholesterol IS present in the membrane of
eukaryotic cell
EXAMPLE
Bacteria, blue green algae and archaebacteria.
Algae, protozoa, fungi, plants and animal«.
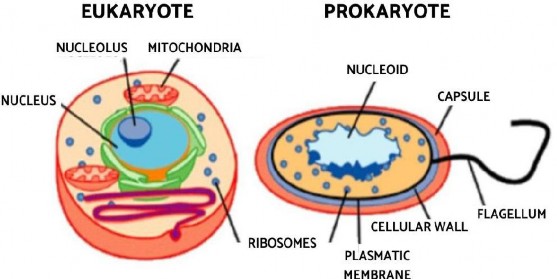
Figure 1.30: A prokaryotic and Eukaryotic cell
: Cell Signaling
Cell signaling is the process by which cells communicate with each other and respond to their environment to coordinate and regulate various cellular activities. It involves the transmission of molecular signals from one cell to another, or from a cell to itself, to elicit a specific response or change in behavior. The basic idea of a signaling pathway can be summarized in a simple diagram (Figure 1.31).
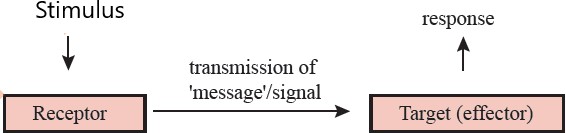
Figure 1.31: Summary of a signalling pathway
Cell signaling can occur over short distances or long distances. It involves ligand-receptor interaction at the target site.
Ligand: A signaling molecule that binds to a specific site on a receptor is called a ligand. After binding, it forms a ligand-receptor complex, initiating a biological response within a cell. By nature, ligand can be proteins, lipids and gases among others.
Receptor: A receptor is a protein molecule usually found on the surface of a cell or within its interior that binds to specific ligands. When a ligand binds to its receptor, it causes the receptor to change its shape, which can activate or inhibit the receptor's associated signalling pathways. Receptors are typically highly selective, binding only to specific ligands or types of ligands. This selectivity is crucial because it ensures that cells respond only to the appropriate signals.
The Three Stages of Cell Signaling.
The process of cell signaling occurs in three main stages: reception, transduction, and response. Reception begins when a ligand binds to a specific receptor, either on the cell surface or within the cell, activating the receptor to initiate the signaling process.
Transduction occurs after the receptor is activated. The signal is relayed and amplified through a series of biochemical reactions inside the cell, ensuring the signal reaches its appropriate targets efficiently. This stage is crucial for accurately transmitting the signal to produce the appropriate cellular response. Finally, in the response stage, the signal triggers a
specific cellular action, such as changes in gene expression, enzyme activity, or cytoskeletal arrangement. This process ensures that vital cellular activities are precisely coordinated and occur within the organism at the right time and place.
Pathway of a Signal
The process by which a cell converts an external signal into a specific cellular response through a series of biochemical interactions is called the signal transduction pathway. Each step in a signal transduction pathway is highly regulated and can involve amplification of the signal, ensuring that even small amounts of a ligand can have significant effects on a cell.
Protein Signal Pathway
Protein signals cannot go through the cell membrane because they are hydrophilic (water- attracting), so they attach to special receptors on the cell's surface. These receptors are typically transmembrane proteins that span across the cell membrane. When a protein signal binds to its matching receptor, the receptor changes shape, getting activated. This activated receptor then start a chain of reactions inside the cell. This chain passes the signal through different molecules within the cell, like cyclic AMP (cAMP), calcium ions (Ca2+) or protein kinases. These molecules help relay the signal to where it needs to go inside the cell. Signal transduction amplifies the initial signal, leading to a larger response within the cell. The' signaling pathway results in a specific cellular response like changes in gene expression, modifications in enzyme activity, cell movement, secretion of substances, or other cellular activities (see Figure 1.32).

Figure 1.32: Protain signaling pathway through membrane receptors
Steroid Signaling Pathway
Steroid hormones, being lipid-soluble (hydrophobic), can diffuse through the cell membrane and enter the cell easily. Once inside the cell, steroid hormones bind to specific intracellular receptors located in the cytoplasm or nucleus. These receptors are often transcription factors. The resulting hormone-receptor complex translocates into the nucleus and interacts with DNA (see Figure I
.33). The hormone-receptor complex binds to specific DNA sequences known as hormone response elements (HREs). This binding either stimulates or inhibits the transcription of specific genes, leading to changes in mRNA production. The altered mRNA levels result in changes in the synthesis of specific proteins. These newly synthesized proteins contribute to the cellular response induced by the steroid hormone. Estrogen, progesterone and testosterone are examples of steroid hormone.
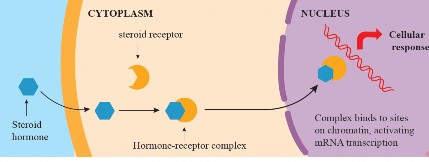
Figure 1.33: Steroid signaling pathway through mobile receptors
Stem cell
Stem cells are undifferentiated cells with the unique ability to develop into various specialized cell types within the body. They have the capacity for self-renewal, enabling them to divide and produce more stem cells, as well as the potential to differentiate into specific cell types with specialized functions (see Figure I .34).

Figure 1.34: Stem cells
Advantages of Stem Cells
Stem cells offer many advantages in various aspects;
Stem cells can replace or repair damaged tissues and organs, offering hope for treating a wide range of diseases and injuries such as Parkinson's disease, type I diabetes, and heart disease.
Stem cells provide a valuable tool for studying developmental and disease processes mechanisms.
They allow scientists to observe how cells differentiate and mature, providing insights into normal development and disease progression. This understanding can lead to breakthroughs in predicting and possibly preventing developmental anomalies.
Stem cells can be used in drug testing and screening to evaluate the safety and effectiveness of new drugs. This can streamline the process of drug development, making it faster and less costly.
Types of Stem Cells

Understanding the categorization of stem cells is important for studying their capabilities and applications in biology and medicine. Stem cells can be broadly categorized based on their origin and their potential to differentiate into other cell types (potency). Based on their potency, students have studied their types in grade 9. Here are the main types of stem cells based on their origin: Embryonic Stem Cells (ESCs): These cells are derived from the inner cell mass of the blastocyst I (early-stage embryo). They are pluripotent and can give rise to nearly all cell types in the body. Their use is often controversial due to ethical concerns regarding the source of embryonic tissue.
Figure 1.35: Human embryonic stem cells
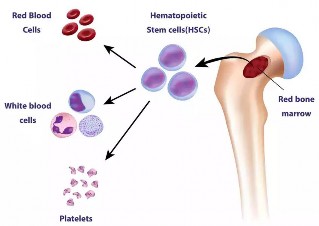
Adult Stem Cells (ASCs): These cells are found in various tissues or organs like the bone marrow, brain, and liver in the body and are also known as somatic stem cells. These cells are generally multipotent, capable of differentiating into a limited range of cell types related to their tissue of origin. They are used to maintain and repair the tissue in which they are found. Common examples include hematopoietic stem cells (found in bone marrow) and mesenchymal stem cells (in bone marrow, adipose tissue, etc.).
Figure 1.36: Adult stem cells in bone marrow
Induced Pluripotent Stem Cells (iPSCs): These are cells that have been genetically reprogrammed to an embryonic stem cell-like state by being forced to express genes important for maintaining the defining properties of embryonic stem cells. iPSCs can be derived from adult cells and are pluripotent, being capable of differentiating into various cell types. They offer a promising source of stem cells without the ethical issues associated with ESCs.
Induced pluripotent stem cells (iPSCs) have revolutionized the field of regenerative medicine and research due to their unique properties. Here are the advantages and disadvantages associated with their use.
Advantages of Induced Pluripotent Stem Cells:
There are many advantages of using induced pluripotent stem cells; Unlike ESCs, which are derived from embryos, iPSCs avoid the ethical concerns associated with the destmction of human embryos, as they are generated from adult cells such as skin or blood cells.
Since iPSCs can be derived from the patient's own cells, they are genetically identical to the original donor's cells. This personalization allows for minimized risks of immune rejection when used in therapies such as tissue transplants.
iPSCs can be cultured indefinitely in the lab, providing an inexhaustible source of patient- specific cells for medical research or treatment.
iPSCs enable the creation of cell models for various diseases, palticularly those for which there is no adequate animal model. This aspect is crucial for studying disease mechanisms, and progression, and for screening potential therapeutic drugs.
Disadvantages of Induced Pluripotent Stem Cells:
Besides advantages, there are some disadvantages of induced pluripotent stem cells, which are;
One of the significant risks associated with the use of iPSCs is their potential to form tumors. The reprograming process can induce mutations that may increase the risk of cancerous growth if the cells are implanted in patients.'
Sometimes, the reprogramming process is incomplete, resulting in cells that retain some memory of their previous cellular identity. This memory could influence the cells' behaviour and efficiency in differentiating into other cell types.
The production of iPSCs is a complex and time-consuming process that requires sophisticated equipment and high levels of expertise, which can be expensive. This may limit the availability of technologies based on iPSCs, especially in lower-resource settings.
Mitosis and Meiosis
The cell cycle is a complex series of events that cells undergo to divide and produce new cells. This cycle is crucial for growth, development, and repair in multicellular organisms and consists of four main phases: Gl (Gap 1), S (Synthesis), G2 (Gap 2), collectively called Interphase, and M (Mitosis). In Gl, the cell grows and carries out its normal functions. The S phase is where the cell duplicates its DNA, preparing for division. In G2, further growth occurs, and the cell ensures that it is ready for division. Mitosis, the final phase, involves the actual division of the parent cell resulting in two daughter cells. Throughout these phases, various checkpoints ensure the cell is properly prepared to move to the next stage, safeguarding the accuracy of cell division.
Mitosis is a process that allows a single eukaryotic cell to divide and produce two genetically identical daughter cells. This process is essential for growth, tissue repair, and the maintenance of the body's various tissues and organs. Mitosis is responsible for the proliferation of somatic (body) cells and occurs in multicellular organisms, including plants, animals, and fungi.
Mitosis is a continuous cellular process, but it is conventionally divided into two main phases: Karyokinesis, which involves the division of the cell nucleus, and Cytokinesis, which encompasses the division of the entire cell.
Karyokinesis is the process of nuclear division during cell division. It involves the segregation and distribution of genetic material (chromosomes) into daughter nuclei, ensuring that each resulting cell has a complete and identical set of chromosomes.
The mitotic apparatus is important for chromosome movement and separation during mitosis, specifically in karyokinesis. It consists of microtubules, associated proteins, and centrosomes that organize within the cell to ensure accurate chromosome distribution. The main components include:
Spindle Microtubules: Protein filaments made of tubulin that extend from centrosomes at opposite cell ends, forming the spindle framework.
Centrosomes: Regions that organize microtubules. They duplicate and move to opposite poles, giving rise to spindle fibers.
Kinetochore Microtubules: Attach to kinetochores (protein structures) on chromosome centromeres, separating sister chromatids during anaphase.
Non-Kinetochore Microtubules: Extend from spindle poles to the cell center, interacting with microtubules from the opposite pole to push spindle poles apart during anaphase.
Astral Microtubules: Radiate from centrosomes toward the cell cortex (the outer layer of the cytoplasm), helping position the spindle apparatus and orient cell division.
The mitotic apparatus ensures chromosomes are evenly distributed to daughter cells, maintaining genetic integrity and accuracy during cell division.
Prophase: During prophase, several significant events take place as the cell prepares for nuclear division. Firstly, the chromatin, a complex of DNA and proteins, begins to condense, transforming into visible and tightly coiled structures known as chromosomes. Each chromosome appears to have two sister chromatids attached at the centromere, The
nucleoli disappear. Simultaneously, thee nuclear envelope, which separates the nucleus from the rest of the cell, staffs to break down, allowing access to the spindle fibers (see Figure 1.37). These fibers capture the condensed chromosomes by attaching to protein structures called kinetochores at the centromeres. The centrosomes start to migrate to opposite ends of the cell. The centrioles extend a radial array of microtubules towards the nearby plasma membrane when they reach the poles of the cell. This management of microtubules is called an aster.
Prophase sets the stage for subsequent mitotic stages, including metaphase, anaphase, and telophase, where further processes lead to the accurate segregation of genetic material into two daughter cells.
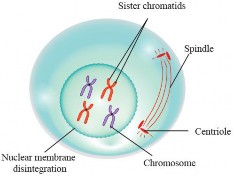
Figure 1.37: Prophase of mitosis
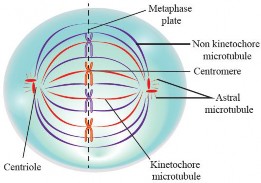
Metaphase: It is a pivotal stage in mitosis where chromosomes align along the cell's center, known as the metaphase plate (see Figure 1.38). This alignment ensures the precise distribution of genetic material to daughter cells. It also serves as a critical checkpoint to confirm the correct chromosomal attachment to the spindle fibers before progressing to anaphase, where sister chromatids are separated.
Figure 1.38: Metaphase of mitosis
Anaphase: It is a crucial stage in mitosis, during which sister chromatids, previously held together at the centromere, are separated and pulled towards opposite poles of the cell (see Figure 1.39). This process ensures that each daughter cell receives an identical and complete set of chromosomes, contributing to genetic stability and the formation of two genetically identical daughter cells. Anaphase is marked by the rapid movement of chromatids along the spindle fibers, and it is a key step in the overall process of cell division.
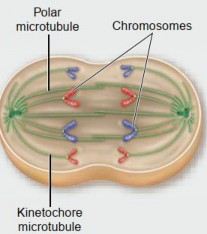
Figure 1.39: Sister chromatids are separating during anaphase of mitosis
Telophase: It marks the near conclusion of mitosis, playing a vital role in the process of cell division. During this stage, the sister chromatids, which were separated and pulled towards opposite poles during anaphase, begin to decondense, transitioning from visible chromosomes back to their more relaxed chromatin form (see Figure 1.40).
Simultaneously, the cell starts to reform a new nuclear envelope around each set of chromatids, establishing two separate nuclei within the cell.

Figure 1.40: Telophase of mitosis
During the latter stages (of telophase, astral microtubules convey signals to the central region of the cell. These signals initiate the activation of actin and myosin proteins, which come together to construct a contractile ring. Over time, this ring progressively contracts, giving rise to the development of a cleavage furrow that becomes more pronounced toward the center of the cell (see Figure 1.41). Ultimately, this sequence of events leads to the division of the original cell into two distinct daughter cells.
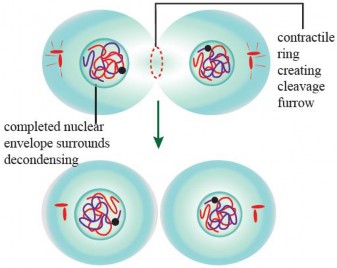
Figure 1.41: Cytokinesis in an animal cell
However, due to the cell wall in plant cells, the phragmoplast, a structure made of microtubules and proteins, directs the formation of the cell plate from Golgi vesicles, which gradually develops into the new cell wall that split the cell into two new cells (see Figure 1.42).
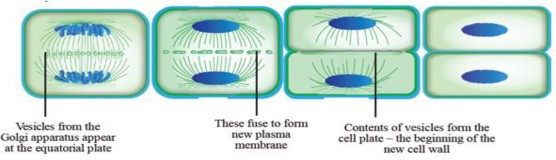
Figure 1.42: Cytokinesis in a plant cell
Meiosis is a unique form of cell division that halves the chromosome number in daughter cells compared to the parent cell. Meiosis exclusively occurs in diploid cells, taking place during gamete formation in animals and spore production in plants.
Following meiosis, each diploid cell generates four haploid cells, a process achieved through two consecutive divisions after a single DNA replication event. These divisions are referred to as meiosis I and meiosis II. The initial meiotic division, known as the reduction division, is responsible for reducing the chromosome count, while the second meiotic division closely resembles the process of mitosis.
Prophase I: Prophase I is a critical stage within meiosis I, which is the first round of meiotic cell division. It is a complex process with several distinct sub-stages, each contributing to the overall genetic diversity of offspring.
Prophase I consists of the following stages:
Here are the stages of Prophase I (also see Figure 1.43):

Figure 1.43: Stages of Prophase
Leptotene: Chromatin within the nucleus begins to condense, and chromosomes become visible under a microscope. Each chromosome consists of two sister chromatids due to DNA replication in the preceding interphase.
Zygotene: During this stage, homologous chromosomes start to pair up through a process called synapsis. This pairing results in the formation of a structure known as a tetrad or bivalent, which comprises four chromatids (two from each homologous chromosome).
Pachytene: In this stage, the tetrads continue to condense and become more tightly aligned. A significant event in pachytene is crossing over, where genetic material is exchanged between non-sister chromatids of homologous chromosomes. This genetic recombination enhances genetic diversity.
Diplotene: During diplotene, the paired homologous chromosomes begin to separate slightly, although they are still connected at the chiasmata (the sites where crossing over occurred). The chromosomes remain attached by these crossover points until the later stages of meiosis.
Diakinesis: In the final stage of prophase I, the tetrads complete their condensation, and the nuclear envelope begins to break down. Chromosomes become fully visible, and spindle fibers start to interact with the condensed chromosomes, preparing for v their eventual separation. Finally, nucleoli disappear.
Metaphase I: Before meiosis I, the homologous chromosomes (pairs of chromosomes, one from each parent) has already undergone replication in the S-phase of interphase I. The homologous chromosomes align themselves along the metaphase plate (also called the equatorial plate) at the center of the cell. Each homologous pair lines up side by side,
with one chromosome from the maternal parent and one from the paternal parent (see Figure 1.44). Spindle fibers attach to the centromere of chromosomes. Each chromosome of a homologous pair receives spindle fibers from one pole.

Figure 1.44: Event of metaphase I of meiosis I
Anaphase I: Anaphase I begins with the separation of homologous chromosomes. Each homologous pair is pulled apart and moved toward opposite poles of the cell (see Figure 1.45). This separation is accomplished by the contraction of microtubules that make up the spindle fibers attached to the centromeres of the chromosomes. As the homologous chromosomes o move to opposite poles, the chromosome number in each daughter cell is reduced by half This is a crucial step in meiosis because it ensures that the resulting daughter cells will have the haploid number of chromosomes, which is half the diploid number. In humans, the haploid number is 23, while the diploid number is 46.
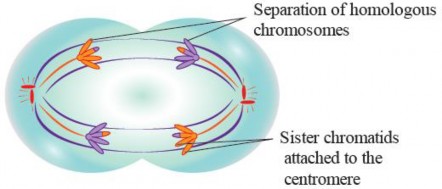
Figure 1.45: Anaphase of meiosis I
Telophase I: During Telophase I, the separated homologous chromosomes, which were pulled apart during Anaphase I, continue to move towards opposite poles of the cell. This movement is facilitated by the contraction of microtubules that make up the spindle fibers. As the homologous chromosomes reach their respective poles, the nuclear envelopes or membranes that had disintegrated during an earlier stage of meiosis (Metaphase I) start to reform around each set of chromosomes. This reformation marks the establishment of two nuclei in a cell. The chromosomes, which were highly condensed and visible during earlier stages, begin to decondense as they are enclosed within the newly formed nuclei. They become less compact and return to their extended, interphase-like state. Telophase I is followed by cytokinesis, which is the division of the cytoplasm and other cell organelles. In animal cells, a cleavage furrow forms, pinching the cell membrane and ultimately resulting in the separation of the two daughter cells. In plant cells, a cell plate directed by phragmoplast forms in the middle of the cell and develops into a new cell wall, leading to the formation of two distinct daughter cells.
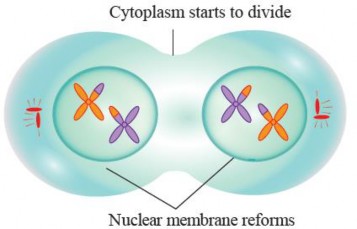
Figure 1.46: Telophase I
Meiosis II
After meiosis I, two daughter cells go through a brief interphase, but unlike the interphase of mitosis, there is no replication of chromosomes. Prophase II, metaphase II, anaphase II, and telophase II closely resemble the respective phases of mitosis. During these stages, chromosomes condense, the mitotic apparatus forms, chromosomes align at the equator, individual or sister chromatids separate, and ultimately, four nuclei are formed at the respective poles of the two daughter cells formed after meiosis I (see Figure 1.47). Cytokinesis occurs, resulting in the formation of four haploid cells, each with half the number of chromosomes (chromatids).

Figure 1.47: Stages of meiosis I and II